
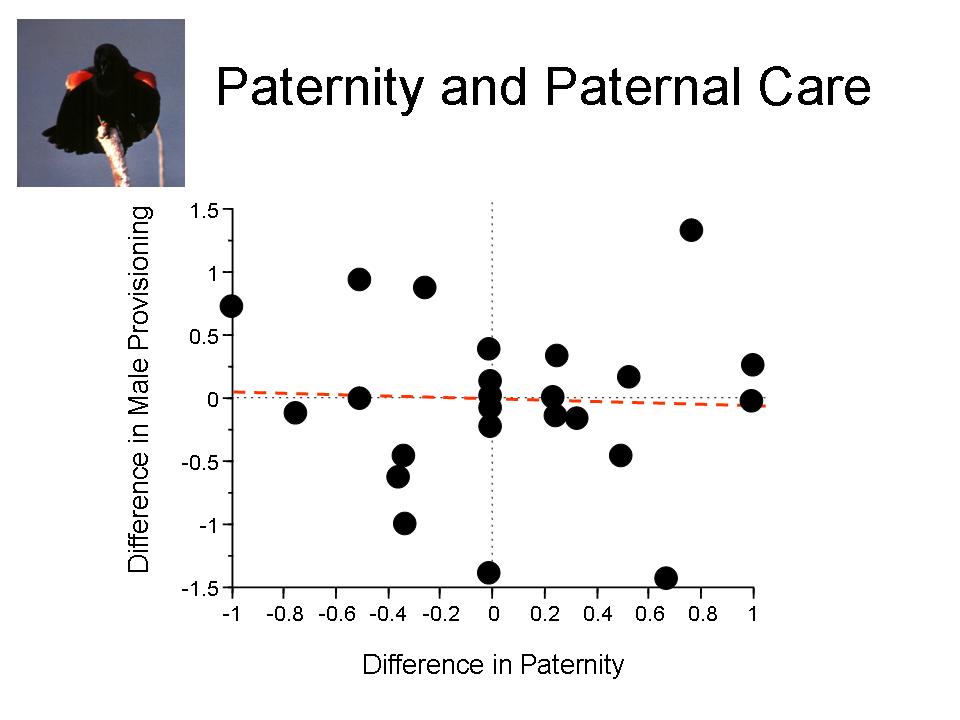
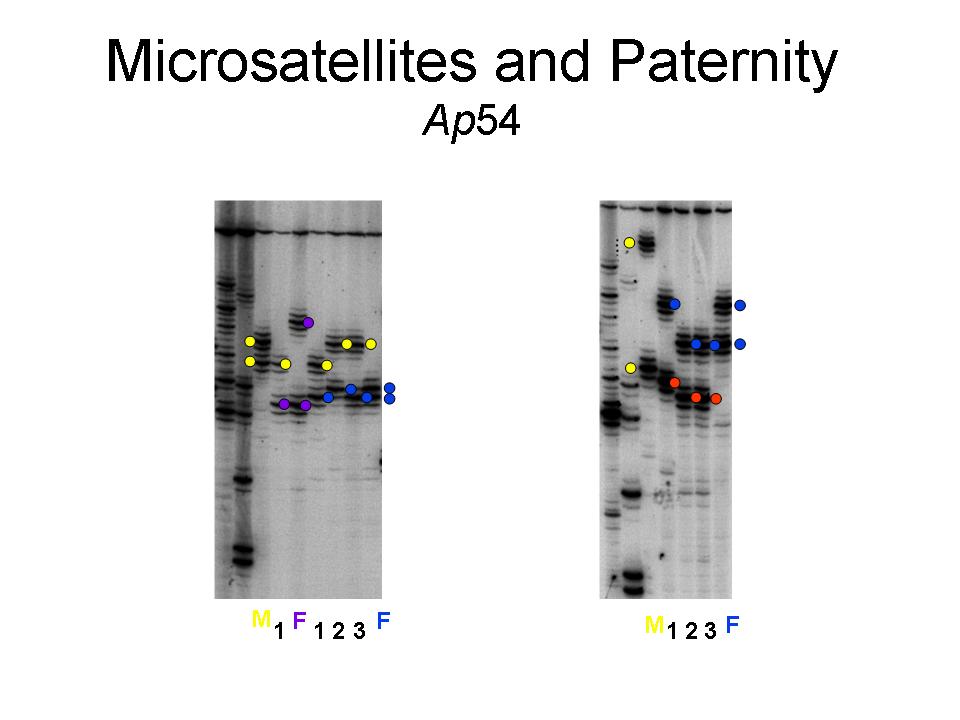
(D) Does paternity influence paternal behavior? I have approached this question both theoretically and empirically. The answers are not simple, and the data suggest some fascinating complexities. Male red-winged blackbirds do not provide food to nestlings in relation to their paternity. The circumstances surrounding the patterns of paternity in the population and the means by which extra-pair copulations occur and lead to fertilizations suggests that red-winged blackbirds fit those situations in which paternity should have relatively little effect on paternal behavior. For more information, see Westneat and Sherman 1993, Westneat 1995, Westneat et al. 1995, Westneat 2001).

These data are currently being analyzed and have yet to be written up for publication.

(G) Several undergraduates and I (in collaboration with Tim Birkhead's lab in Sheffield, UK) examined male sperm counts using model females (Westneat et al. 1998). We found that male ejaculate size decreased with multiple copulations in a short period of time, but appeared to recover after a delay of an hour between copulations.
(H) Recently, we have surveyed the microbes present in male ejaculates and in the cloacae of males and females. The goal was to uncover the possible importance of sexual transmission of fitness-affecting bacteria (see Westneat and Rambo 2000 or Hupton et al. 2003). We found bacteria in many male ejaculates but no one type predominated, meaning that every male had a unique assemblage of types. This has important implications for understanding the risks of multiple mating for females and for further studies of sexually transmitted microbes.
(I) In collaboration with Dennis Hasselquist of Lund University, Sweden, we have used Diphtheria-Tetanus antigens to measure the responses of the male immune system to presumably novel antigens. John Wingfield of the University of Washington analyzed testosterone titer for these birds as well. We found few significant relationships (Westneat et al. 2003), which complicates our understanding of the potential links between advertisements, immunity, and hormones. See also Westneat and Birkhead 1998 for some background ideas of relevance to this project.
Acknowledgments
The red-winged blackbird project has benefited from the efforts of a very large group of people. Funding has been provided by the National Science Foundation, The University of Kentucky, and Cornell University.
Ithaca Study Site
Cornell University Pond Unit, Bob Johnson, Dr.
Nelson Hairston, Jr., Dr. Ann Clark and her field crew, and field assistance
from Christine Butler, Cathy Corson, Dan Christian, Karin Gastreich, Jennifer
Davis, Jeff Dickhaut, and Herman Mays.
Kentucky Study Site
Peabody Mining Co., KY Fish and Wildlife Resources, and field help from Tony Roberts, Zeb Weese, Emily Reed, Maribeth Polhill, Lisa McGraw, Pete Blank, Chad Johnson, Tasha Macilveen, Sarah Laughlin, Jennifer Fraterrigo, Michelle Brant, Nevelyn Russell, Brian Smith, Jamie Moseley, Jessica Brock, and Matthew Shawkey.
Lab
Thanks to Dr. Chip Aquadro at Cornell for his support during the early part of this project. Assisting on the paternity analyses in the lab were Lorna Breen, Greta Crutcher, Tamara Roush, Brent Little, Jana Hawks, Jamie Moseley, Richard Hanschu, and Herman Mays.