
Sabbatical at Seoul National Univ.
2006 go to www page
| LATEST RESEARCH | ||||||
| Back to General Research | ||||||
| Back to Home Page | | |||||
Sabbatical at Seoul National Univ. 2006 go to www page | ||||||
|
Three main current projects: 1. Social interaction among crayfish of same and difference species 2.
Effects of bacterial endotoxin lipopolysaccharide on cardiac function
and synaptic transmission |
||||||
|
1. Social interaction among crayfish of same and different species |
||||||
| Go to web page... (go to) | ||||||
| 2. Effects of bacterial endotoxin lipopolysaccharide on cardiac function and synaptic transmission | ||||||
|
Cooper, R.L., McNabb, M. and Nadolski, J. (2019) The effects of a bacterial endotoxin LPS on synaptic transmission at the neuromuscular junction. Heliyon- Elsevier, 5 (2019) e01430 https://www.heliyon.com/article/e01430 Anyagaligbo, O.,
Bernard, J., Greenhalgh, A. and Cooper, R.L. (2019)
The effects of bacterial endotoxin ( LPS ) on cardiac function in
a medicinal blow fly (Phaenicia sericata) and a fruit fly (Drosophila
melanogaster). Comparative Biochemistry and
Physiology- Part C 217:15-24. 5 year impact factor 2.819. [PDF] Saelinger, C.M., Danley, M. and Cooper, R.L. (2019). The effects of bacterial endotoxin (LPS) on synaptic transmission at neuromuscular junction in an amphibian. International Journal of Zoological Research 15(2):38-42. https://scialert.net/current.php?issn=1811-9778 [PDF] Saelinger, C.M., McNabb, M.C., McNair, R., Bierbower, S. and Cooper, R.L. (2019) Effects of bacterial endotoxin (LPS) on the cardiac function, neuromuscular transmission and sensory-CNS-motor nerve circuit: A crustacean model Comparative Biochemistry and Physiology, Part A. 237:110557. 5 year impact factor 2.173 https://www.sciencedirect.com/science/article/pii/S1095643319303216 [ PDF ] https://doi.org/10.1016/j.cbpa.2019.110557 Ballinger-Boone, C., Anyagaligbo, O., Bernard, J., Bierbower, S.M., Dupont-Versteegden, E.E., Ghoweri, A., Greenhalgh, A., Harrison, D., Istas, O., McNabb, M., Saelinger, C., Stanback, A., Stanback, M., Thibault, O., and Cooper, R.L. (2020) The effects of bacterial endotoxin (LPS) on cardiac and synaptic function in various animal models: Larval Drosophila, crayfish, crab, and rodent. International Journal of Zoological Research 16: 33-62. doi: 10.3923/ijzr.2020.33.62https://scialert.net/abstract/?doi=ijzr.2020.33.62...[PDF] Istas,
O., Greenhalgh, A. and Cooper, R.L. (2020)
Repetitive exposure to bacterial endotoxin LPS alters synaptic transmission.
Journal of Pharmacology and Toxicology 15:
65-72. DOI: 10.3923/jpt.2020.65.72http://docsdrive.com/pdfs/academicjournals/jpt/2020/65-72.pdf Bernard, J., Marguerite, N., Inks, M. and Cooper, R.L. (2020) Opposing responses of bacterial endotoxin lipopolysaccharide (LPS) and TRPA1 thermal receptors on synaptic transmission and resting membrane potential. Current Research in Bacteriology 13 (1):10-21. DOI: 10.3923/crb.2020.10.21 http://docsdrive.com/pdfs/ansinet/crb/2020/10-21.pdf |
||||||
| ABSTRACT
Serotonin (5-HT) levels
in the hemolymph of crustaceans is implied to alter aggressiveness which
impacts social interactions. The activation of IP3 as a second messenger
cascade within crayfish motor neurons in response to application of 5-HT,
suggests that the 5-HT receptor subtypes on the motor neurons are analogous
to the vertebrate 5-HT2A receptors. Based on evidence in other systems,
it would be expected that chronically sustained 5-HT levels of aggressive
individuals would result in a compensatory negative feed-back regulation
and/or that target tissues would diminish their sensitivity to high levels
of circulating, free 5-HT. We addressed the issue of up- and down-regulation
in the sensitivity of the responsiveness to exogenously applied 5-HT at
the NMJs of crayfish in which the animals have altered endogenous 5-HT
levels. Injections of the 5-HT1 and 5-HT2 vertebrate receptor agonist,
m-CPP, for 1 week resulted in a decreased responsiveness to application
of 5-HT. The compound p-CPA blocks the enzymatic synthesis of 5-HT and
following 7days of p-CPA injections, a super-sensitivity to exogenous
application of 5-HT for both tonic and phasic NMJs was observed. However,
acute applications of p-CPA and m-CPP, followed by extensive saline washing,
did not reveal any altered receptivity to 5-HT application. |
||||||
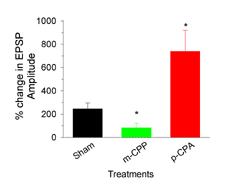 |
||||||
| Chronic exposure to m-CPP or p-CPA altered the synaptic responsiveness of the neuromuscular junctions to 5-HT. Trains of 40 Hz containing 10 events were induced every 10 seconds. The amplitude of the 10th EPSP responses were measured from the trough to the peak before and after 5-HT (100nM). The mean (+/- SEM) of the percent differences for the sham (vehicle), m-CPP, and p-CPP groups (n=6) are shown. The star denotes a significance (P<0.05; Mann-Whitney non-parametric test) compared to the sham group. | ||||||
| This
project in regulation of receptor sensitivity for neuromodulators is a project
primarily conducted by undergradutes. It is well established that receptors undergo up- and down-regulation via alteration of their expression levels and/or densities on cell surfaces (Azaryan et al., 1998). Altered cellular activity as well as the action of agonists or antagonists being bound to a receptor can induce regulation in the levels of functional receptors (Kokay and Mercer, 1997; Welner et al., 1989; Fone et al., 1998). For example it has been shown that 5-HT1A receptors will demonstrate desensitization when either an agonist or antagonist is present (Hensler and Truett, 1998). Even naturally induced down regulation of 5-HT2c receptors can be induced as a result of exercise (Broocks et al., 1999). The 5-HT1 and 5-HT2 receptor agonist 1-(3- Chlorophenyl) piperazine dihydrochloride (m-CPP) has been observed in rats to down regulate receptor numbers (Fone, et al., 1998). The precise mechanism of action in regulation of the 5-HT receptors has not yet been elucidated. The purpose of this project is to address the issue of up- and down-regulation of the sensitivity of responsiveness to exogenously applied 5-HT at the neuromuscular junctions of crayfish which have reduced endogenous 5-HT levels (enzymatic inhibition of the synthesis of 5-HT) or have chronic presence of an agonist in their hemolymph. Reduction of systemic levels of 5-HT in the crayfish is likely to result in up-regulation of 5-HT receptors since these animals normally contain 5-HT in their hemolymph (open circulatory system). In contrast, chronic high levels of the 5-HT agonist m-CPP, would likely result in a down-regulation of the 5-HT receptors so that exogenous application of 5-HT would show reduced responsiveness as compared to sham injected animals. In preliminary studies, we have demonstrated alterations in the sensitivity to exogenous application of 5-HT at the crayfish neuromuscular junction after altering levels of the endogenous production of 5-HT and exposure to a 5-HT agonist. Future work will be focused at identifying the types of 5-HT receptors present that the crayfish NMJ and the mechanisms behind the physiological responses that we have observed in sensitivity to 5-HT. (Recent progress was presented at the SICB meeting in Chicago, IL in January 2001 & up-dates will be presented at this year's Neuroscience Meeting. Multiple research disciplines are being examined separately but they are all interrelated and focus on the issue of synaptic function. With the successful outcome of findings over the last 4 years I believe this approach will continue to bear fruit.
Azaryan, A.V., Clock, B.J., Rosenberger, J.G., and Cox, B.M. (1998) Transient upregulation of mu opioid receptor mRNA levels in nucleus accumbens during chronic cocaine administration. Can. J. Physiol. Pharmacol.76:278-283. Broocks,
A., Meyer, T., George, A., Hillmer, Vogel, U., Meyer, D., Bandelow, B., Hajak,
G., Bartmann, U., Gleiter, C.H., and Ruther, E. (1998) Decreased neuroendocrine
responses to meta-chlorophenylpiperazine (m-CPP) but normal responses to ipsapirone
in marathon runners. Can. J. Physiol. Pharmacol. 76:278-283. Dixon,
D. and Atwood, H.L. (1989) Conjoint action of phosphoinositol and adenylate cyclase
systems in serotonin-induced facilitation at the crayfish neuromuscular junction.
J. Neurophysiol. 62, 1251-1259. Fone, K.C., Austin, R.H., Topham, I.A.,
Kennett, G.A., Punhani,T. (1998) Effect of chronic m-CPP on locomotion, hypophagia,
plasma corticosterone and 5-HT2C receptor levels in the rat. British J. Pharmacol.123:1707-1715.
Hensler, J.G. and Truett, K.A. (1998) Effect of chronic serotonin2
receptor agonist or antagonist administration on serotonin 1A receptor sensitivity.
Neuropsychopharmacology 19:354-64 Kokay, I.C. and Mercer, A.R. (1997) Age-related changes in dopamine receptor densities in the brain of the honey bee, Apis mellifera. J. Comp. Biochem.181:415-423. Welner, S.A., De Montigny,
C., Desroches, J., Dejardins, P., and Suranyi-Cadotte, B.E. (1989) Autoradiographic
quantification of serotonin 1A receptors in rat brain following antidepressant
drug treatment. Synapse 4:347-352. |